After just over 6 years on Blogger, Pseudoplocephalus is moving to a new home at www.pseudoplocephalus.com! It's finally time for me to have a proper grown-up website for responsible adults, so you'll be able to find not only the blog, but information about my research, publications, teaching, fieldwork, science communication, and public science efforts all in one place. If you aren't already following me on Twitter (@VictoriaArbour), you can also do that too! Thank you, dear readers, for following along all of these years, and I hope I'll see you over at my new digs.
Friday, April 15, 2016
Wednesday, March 9, 2016
Ankylosaurs by the sea
Ankylosaurs, like probably most other dinosaurs, were
landlubbing, terrestrial animals without obvious aquatic adaptations. And yet,
surprisingly, their fossils are found in marine sedimentary environments more
often than most other dinosaurs (except hadrosaurs). Some, like Aletopelta,
wound up in shallow or lagoonal environments - Aletopelta's carcass became a
reef! - but some, like the Suncor nodosaurid, wound up far away from shore.

Aletopelta! See if you can spot the oyster marks, invertebrates, and shark teeth around the pelvis and legs.
I wanted to look into this phenomenon in a bit
more detail than it had been investigated previously, and with my work on
ankylosaurid phylogenetics and biogeography published last year the time was right
to dive into marine ankylosaurs. I added in more taxa into my ankylosaurid
character matrix to check whether there were any secret ankylosaurids hiding in
the taxa I didn't previously include.
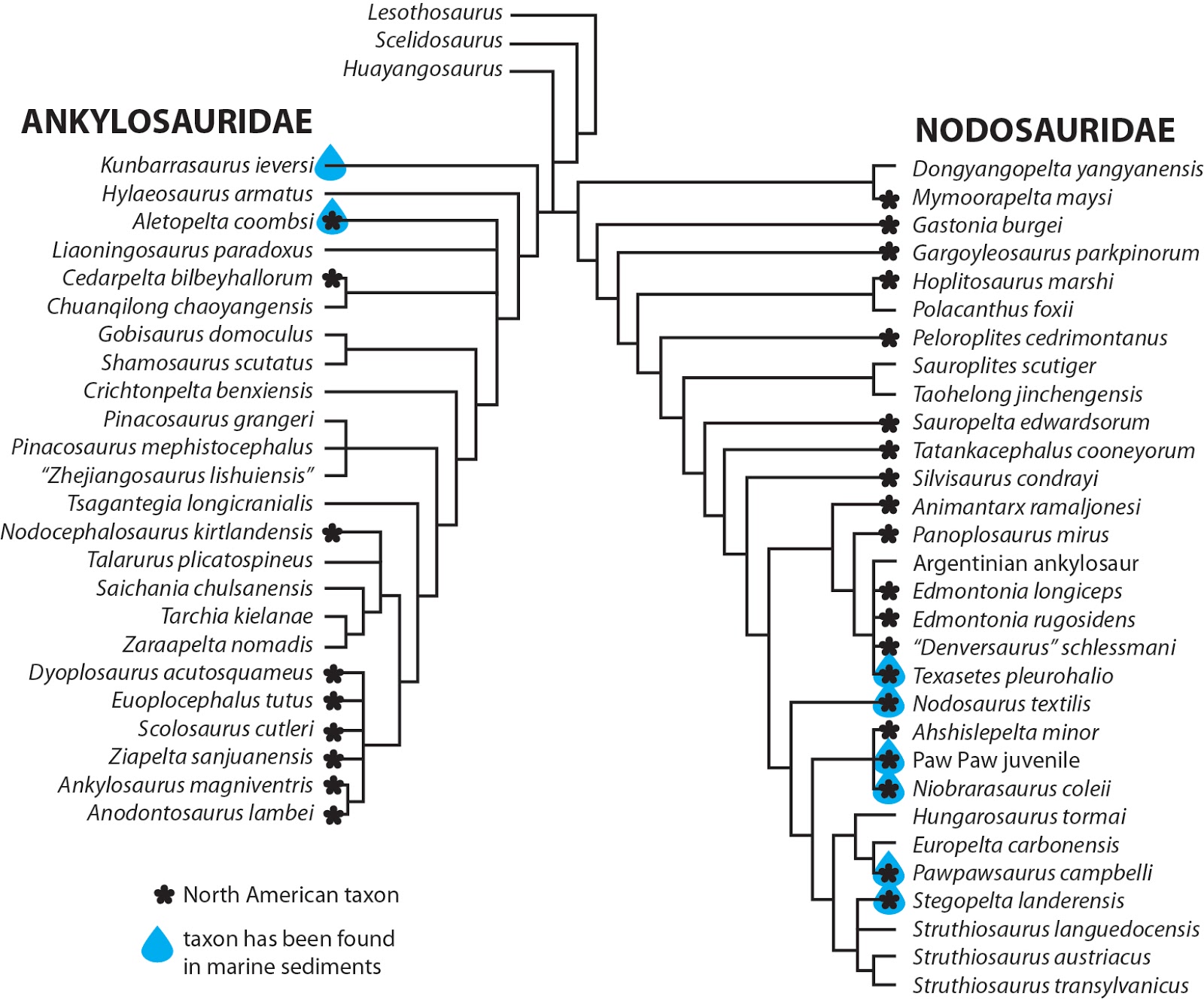
Surprisingly, Hylaeosaurus popped out as
a basal ankylosaurid in this analysis, an intriguing result that I think needs
further investigation but could be really interesting. Mostly everything else that should be a nodosaurid was a nodosaurid, and I got surprisingly good resolution in the Nodosauridae, but take the interrelationships there with a tiny grain of salt given that I didn't make any new characters for this analysis and the existing character set is geared towards ankylosaurids.
Next up, I downloaded a dataset of all the ankylosaur
occurrences in the Paleobiology Database and went through each entry (more than
500!) to verify whether or not the specimens were ankylosaurids, nodosaurids,
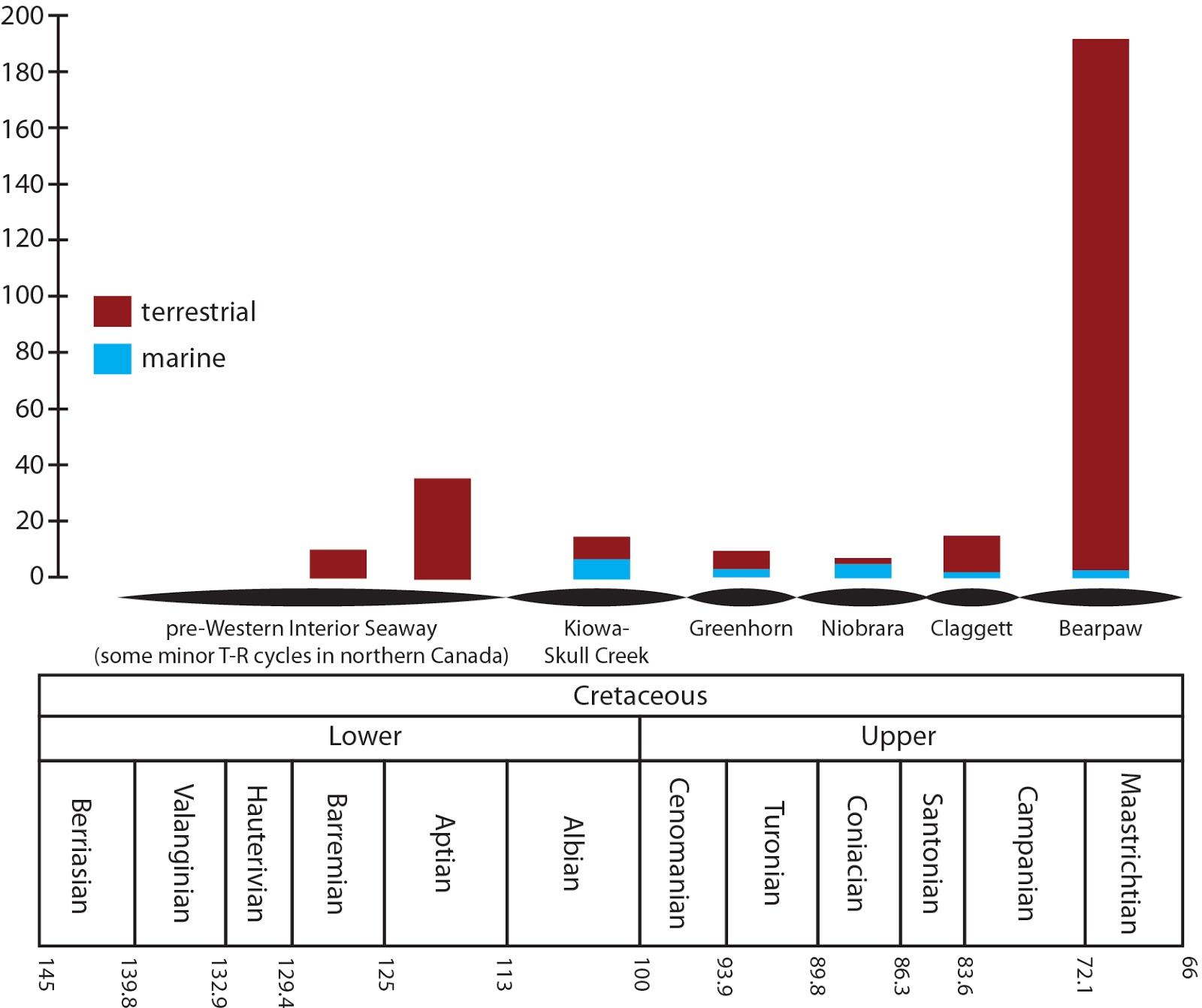
or if we couldn't tell for sure, and their depositional environment. With that data in hand, we wanted to know what the
geographic distribution of ankylosaur marine occurrences looked like, and here
it is:

Lots of ankylosaurs in the northern hemisphere, not so many
in the southern hemisphere. More than half of marine ankylosaurs occur in North
America, which is perhaps unsurprising given that North America had a bad habit
of being underwater for a lot of the Cretaceous. When we exclude indeterminate
occurrences, we get an ankylosaurid-nodosaurid marine-terrestrial split that
looks like this:

In North America and Asia, the distribution of ankylosaurs in different environments is statistically significant, but it isn't in Europe (and remember, Hylaeosaurus hasn't previously been found as an ankylosaurid so there might not even be ankylosaurids in Europe).

Sunday, February 7, 2016
Superb Lemur Sunday
On this most hallowed of Super Bowl Sundays, I watched some lemurs instead of football.
Lemurs are very busy at all times!
The paleo crew from the museum and NCSU decided to visit the Duke Lemur Center, a research facility at Duke University about 30 minutes from Raleigh with the largest and most diverse collection of lemurs outside of Madagascar. Although it is primarily a scientific research facility, the DLC also offers tours and educational workshops.

It's a large facility with numerous enclosures for the different species they keep, including these 'summer homes' for when the weather is a bit warmer. The DLC also has multiple large multi-acre forest habitats where the lemurs can freely roam during the daytime in the summer. I'd love to go back for one of the forest tours!

At the beginning of the tour there's an opportunity to check out some cool anatomical specimens, like the hand and skull of the insectivorous aye-aye! Check out that thin probing finger!

And you can also touch some pelts from some of the individuals that died of natural causes. I can confirm that lemurs are extremely soft. Also of interest is the wide variety of colours and patterns that lemurs have - I love the little white blaze on the top of the head of the red ruffed lemur (Varecia rubra), the pelt in the centre of this photo.

And there were plenty of live lemurs for us to meet as well! One really cool thing about the DLC is that there are lemurs that are not found in most zoos because they are either relatively delicate in captivity, or destructive of their habitats and require a lot of maintenance. Coquerel's sifakas (pronounced "shifawkas"; Propithecus coquereli) might be familiar to many from the TV show Zoboomafoo! In fact, the show was partly filmed at the DLC.

I also really liked the black and white ruffed lemurs (Variecia variegata).

New to me was the mongoose lemur (Eulemur mongoz), a relatively relaxed fellow compared to the others.

And also new to me were the blue-eyed black lemurs (Eulemur flavifrons). Although it might look like there are two species in the photo below, both lemurs are E. flavifrons! Several lemur species exhibit sexual dichromatism, where males and females are markedly different colours. In this case, the males are black and the females are golden. I'd love to learn what drives this difference in colours, especially when the males aren't particularly elaborate or showy! It's a weird manifestation of sexual selection.

In addition to these lemurs we also got to see some ring tailed lemurs (Lemur catta), which I'm sure mostly everyone is familiar with, a slow loris (Nycticebus coucang), an unfortunate victim of the pet trade despite being toxic, the diminutive gray mouse lemur (Microcebus murinus), and a fat-tailed dwarf lemur (Cheirogaleus medius), which was true to its name.
But the real showstopper for me was a chance to see the remarkable aye-aye (Daubentonia madagascariensis)! Luckily for us, the nocturnal species are kept on a flipped daylight schedule, so we got to see an active aye-aye under dim red light.

Aye-ayes are one of the species that is particularly hard to keep because, as insectivores that gnaw into wood in order to find food, they are rather destructive to their habitats. The DLC is the only facility in the world that has successfully bred aye-ayes, and these are not animals you're going to see at most zoos so it was a real treat to watch them in action. Things I did not realize about aye-ayes: they are big, and they have incredibly fluffy tails! I'd say the aye-aye was about the size of a large house cat, which was much bigger than I had imagined - I think I had always considered them to be one of the smaller species, perhaps squirrel-sized.
The DLC is a pretty cool place doing a lot of interesting conservation work in addition to non-invasive research on lemurs, and if you're in the Raleigh area it's definitely worth a tour!
Monday, February 1, 2016
Cool stuff at my museum.
Today I wanted to share some of the cool stuff the North Carolina Museum of Natural Sciences does besides palaeontology!

My home is in the Nature Research Center, the new addition to the museum that opened in 2012 and which features a 3-story-tall globe on the outside! The globe is always an attention-getter, and most days when I'm walking to the bus there are people outside posing in front for pictures. It's a cool thing to have as the icon for the museum. It looks cool in the rare snow we get here, and moody at night.

But the globe isn't just a cool external feature, it's also an important part of the inside of the museum, as the Daily Planet Theatre. The inside of the globe forms a curved surface visible from all three floors, during which scenes from nature are projected for most of the day. Each day one of the scientist gives a special presentation on their research in this venue, and special guests speak here for events. This is where Lindsay and Susan debuted the Carolina Butcher last spring!

The first floor has this spectacular right whale skeleton, but it's not just any right whale - this one has an important scientific story to tell. If you look closely on the right mandible (on the left in this image), and towards the back, you will see a series of little circular divots. This whale's mandible was sampled in order to understand breaking stresses in whale jaws during ship strikes, the cause of death for this poor fellow. Research on this specific whale helped create new legislation for ship speeds in whale zones. I think it's an awesome example of real science making tangible benefits. It's a great way of linking research to a specimen and giving it a powerful narrative.

Within the Nature Research Center are five active research laboratories - Paleontology, Astronomy & Astrophysics, Genomics & Microbiology, Evolution, and Biodiversity. Our labs are part of the exhibit space and visitors to the museum can peer into the labs. Most of the labs have interesting displays in the windows and one of my favourites is the Biodiversity Lab's dermestid colony. They have bear skulls, deer skulls, coyote bits, and all kinds of things decomposing in full view, and at the moment there is a whole baby seal in there.

Not all of the animals are dead! The museum has a pretty extensive collection of live animals on exhibit, like this matamata turtle. Another really cool thing is the visible veterinary lab in the Nature Research Center, where you can see veterinarians caring for the museum's animals. And sometimes this includes surgery! There is always a bi crowd of people whenever the vet window is occupied. Most recently, the museum took in several cold-stunned sea turtles to rehabilitate.

One of my very favourite things in the whole museum besides the Paleo Lab is the Naturalist Center, basically a hands-on library of STUFF. All kinds of good stuff: bones, fur, feathers, specimens in plastic, stuff to look at under the microscope, field guides and anatomical diagrams. I've never really seen something quite like this in any other museum. It's almost always busy, especially on the late nights on Thursdays, and kids LOVE IT and I think parents probably absorb some information too because everything's hands on.

The thing that gets the most attention is this awesome interactive table where you can take specimens from a specially designated shelf, stick them on, and then a computer somewhere recognizes the specimen and projects a whole bunch of interactive information onto the table. There is much swooping and swishing and it's one of the only touch screen info things I've seen people really use for more than 2 seconds. It's slick, it's cool, and you move a real object around in order to make it work. It's perfect. Here it is in action:
There's so much more that I can't possibly put everything in one post, but I hope this gives you a sense for the unique and interesting ways the museum incorporates real-time scientific research, hands-on and experiential learning, and cool narratives into its exhibits. Things are blended really well and it's no surprise that the museum is one of the most popular destinations in the region. Here's a dinosaur, thanks for sticking around!

Wednesday, January 6, 2016
Social engagement in Dino 101
Happy new year folks! There are no pictures in this post, SORRY NOT SORRY.
A group of researchers at the University of Alberta recently
published a study on learner engagement in Dino 101, and I thought I'd
summarize it briefly here and share a few thoughts about it. You can read the
original article online for free via Google Books: "Emotional and social engagement in a massive open online course: an examination of Dino 101".
You might also want to check out another summary of their data at the
University of Alberta's site.
Daniels et al. lay out four components for describing the
otherwise somewhat nebulous term 'engagement':
- Cognitive engagement (investment into thinking about tasks and mastering content; could also be thought of as motivation for learning)
- Behavioural engagement (things like attendance, paying attention, and participation)
- Emotional engagement (things like anxiety, boredom, interest, etc.)
- Social engagement (willingness to socialize with others, to make connections)
They
mainly investigated emotional and social engagement, since cognitive and
behavioural engagement are largely measured by things like course completion
rates and grade averages. Daniels et al. sent a survey to people enrolled in
Dino101 towards the end of its initial offering, as well as the students
enrolled for credit in Paleo200/201 at the UofA, and also interviewed a total
of 30 students representing all three versions of the course. Because all
of the lessons and study material for Dino101 was released at the beginning of
the semester (in contrast to many MOOCs which release new lessons on a weekly
basis, probably in line with the in-person lectures at the university), I
wonder if conducting the survey at the end of the semester, in December,
influenced the results. We heard from many students that they completed Dino101
very very quickly – some in as little as a day or two, and many within 2-6
weeks. From managing the forums, I can also attest that discussion board
participation dropped pretty dramatically after about six weeks. Therefore,
students that completed the course are not necessarily students that stuck
around until December, and I wonder how well this slice of the long statistical
tail might represent the majority of the people who engaged with Dino101.
Overall,
the results showed that we did a pretty good job of emotionally engaging
Dino101 students: many said they were inspired by the material, didn't get
bored, and felt attentive during the lectures. We fared less well in terms of
social engagement: many students were neutral about whether or not they had a
sense of belonging in Dino101, but only 16% had expected student-student
interactions to contribute to their own learning. (In other words, most
students expected to learn primarily from the videos and course notes, and not
from discussions with other students.) I'm not sure if this is typical for a MOOC or unique to Dino 101, so I'd be curious to see if there are similar studies for other courses that show similar results.
In
the data from the student interviews, Daniels et al. highlighted both positive
and negative aspects to the discussion forums – some students were really
interested in the forums even if they didn't participate, some didn't look at
them at all, and some found them overwhelming because of too many email notices.
I think there are a couple of takeaways just from this one portion of their
paper:
- If you're doing a MOOC, make sure the button for the discussion forums is prominently displayed towards the top of the course page near the lecture videos. I think ours was located kind of far down the menu and some students might just overlook it.
- Give students some guidelines for using the forums, both technically (i.e. how to turn off email notifications for new posts, because that would have driven me COMPLETELY MAD if I hadn't known how to turn it off for most posts), and by giving suggestions for how to participate in the forums. Do this right at the beginning of the course.
Another thing that popped up in this paper is that while social engagement online wasn't
as high as it could be, social engagement in meatspace was unexpectedly common.
There were lots of people taking the course with their kids or other family
members at home, and they would sit and do the course together. So, here is
another takeaway suggestion: find ways to increase social engagement by giving
people things to do at home together, in the physical world. Maybe we could
provide suggestions for local museum trips or fossil sites based on where
people in the course are based, or give some suggestions for hands-on
activities people can do with stuff you find in your house.
Finally, there were mixed feelings about engagement with the
instructors of the course (here limited to Phil Currie and Betsy Kruk, who
presented the material, but excluding myself and the other teaching assistants who
did most of the online interactions in the forums). Some students felt like
they had a good connection with Phil and Betsy despite the scripted presentations,
and others found they didn't like the scripted format at all. Strangely, the
paper doesn't include much discussion about engagement with the instructors (and
here I'm including the TAs) in the discussion forums, the only part of the
course where that was really possible given the pre-recorded nature of the
videos.
My final comment here is that if engagement with an
instructor is important for social engagement in MOOCs (and I think engagement
with the instructor is important in education generally, so it should probably be
important in MOOCs), I don't know what we're going to do. One of the comments
from the interviews that's highlighted in the paper is that one student didn't
have his question answered in the forums – and with my reading between the
lines, that probably means we failed to socially engage this student, which
sucks. I feel badly that a student felt ignored. But the reality is also that
we had 23 000 people enrolled in Dino101, and had to manage hundreds of forum
posts on very limited time budgets, in addition to managing the for-credit
version of the course at the university. There is no way to scale up personal
interactions between students and instructors in a learning environment without scaling up your teaching staff – either you need more instructors (in
the form of profs, TAs, whatever), or you need fewer students, or you're not
going to be able to interact with every student that wants interaction. And we
shouldn't be asking instructors in a university to educate for free, so
somebody needs to be paying for those extra people. So that's one more
important takeaway here: social engagement requires a lot of time investment
from the instructors to encourage discussion and set up an environment that
encourages social interaction, so if you want to run a MOOC with high social
engagement, you need to budget money for lots of instructor/TA time.
Anyway, that's probably one of the last posts I'm going to
have about Dino101/MOOCs for a while, but I thought I would mention one other
piece of news: before I moved down to Raleigh in 2014, I had started work on some
new palaeontology mini-MOOCs in my role as the Science Digital Learning Manager
at the UofA. After much hard work from the palaeo crew at the UofA, these
courses are now just about finished and will be launching on Coursera over the
next few weeks – if you liked Dino101, you might want to take a look at
Theropods and the Origin of Birds (starting later in January), Ancient Marine Reptiles (starting in February), and Early Vertebrate Evolution (starting in
March, not yet available for registration at Coursera but keep an eye on the
UofA's page there).
Tuesday, December 8, 2015
Blog survey results!
It's hard for me to even believe this, but I've been writing
Pseudoplocephalus for over 5 years now. I'd been an avid reader of many science
blogs for a couple of years before I decided I wanted to try it out myself, and
I decided to jump in finally because I was going to be spending three and a
half months working and traveling around Korea, China and Mongolia back in 2010
on an NSERC-funded study abroad kind of thing. I figured that blog updates
would be the best way to show what I was doing to friends and family, and if I
liked it, I'd maybe keep writing about my research afterwards.
As I started writing here, I decided pretty quickly that I
wanted to use Pseudoplocephalus as a science outreach tool (as opposed to
keeping a grad school diary for my own benefits, etc.). At this point, my
general goals with the blog are:
1. Providing summaries of my research papers for nonspecialists,
especially for papers that are paywalled.
2. Showing what it's like to be a research
palaeontologist, for people who aren't scientists. (And, to be visibly female
while doing so.)
3. Promoting the research environments of the
various institutions I've worked at, to help increase the public's appreciation
of research in museums and universities.
4. Talking about other issues of interest to me,
like where palaeontology intersects with popular media and social justice
issues. For this goal, I'm interested in reaching both scientists and
nonscientists.
So, am I accomplishing any of those goals? I took part in
Paige Jarreau's science blog survey and some of you were kind enough to fill
out the survey, so here's a little bit of what I learned and how it relates to
what I'm doing.
How are people finding my blog?
At least in terms of the people who answered the survey,
most people seem to get to my blog via Facebook and Twitter, as well as the
blogrolls of other palaeontology sites. But, about 20% of the respondents had
only read one or two posts on my blog, suggesting that a fair number of people
stumble across it without necessarily being deeply embedded within the
palaeoblogosphere. Most of the survey respondents said they often seek out
information about science online, and nobody said they rarely or never seek it
out. Apparently, several of you were motivated to keep reading because you like
my writing style, so thanks! That is nice of you to say!
How are people using my blog?
Many of the survey respondents read Pseudoplocephalus to
keep up with the latest palaeo news and to find information that might not be
reported in traditional media, so, cool. A bunch of you also wrote in that you
come here to learn about ankylosaurs, so yay! Most of you do not come here for
emotional support, which is also good, because ankylosaurs are terrible
emotional support-givers. I recommend you check out Captain Awkward for the
top-shelf adulting advice. Overall, I'd say the way people seem to be using my
blog is in line with why I'm writing what I write.
Who are you people, anyway?
Several of the survey respondents noted that they know me
personally. I AM BEING WATCHED, THANKS ANONYMOUS FRIENDS FOR CREEPIN' ME OUT.
But seriously, that's not unexpected, and thanks creepy anonymous friends for
filling out my survey. Out of the 102 people who filled out the survey, 67%
identified as male and 31% identified as female. The biggest age cohort is
YOUNGER THAN ME which is giving me SOME KIND OF FEELINGS. Most of you are also
Caucasian, which means I need to do a better job of reaching out to non-white
people or making my blog an inviting space for underrepresented minorities in
science to come and have a look. I am not totally sure how to do this, so I
guess I have some research ahead of me!
I am also apparently in good blog company because lots of
you guys read blogs that I also like to read too, like Tetrapod Zoology,
SV-POW, Laelaps, and Love in the Time of Chasmosaurs. In that same question
(list up to three other science blogs that you read regularly), I noticed that
blogs written by dudes far, far outnumbered blogs written by ladies in the
blogs you submitted. Does this indicate that there are fewer blogs written by
women in the palaeoblogosphere (or at least dinoblogosphere)? Are people more
likely to think more highly of blogs written by men than women, or just more
likely to remember blogs written by men?
Lastly, it was interesting to see the self-identified
occupations you all listed – while it is not surprising that many of you are
grad students, palaeontologists, or other scientists, it was heartening for me
to see that non-scientists are also reading the blog, at least sometimes. Another
goal kind of accomplished!
So, am I meeting any of the general goals I keep in mind
when I'm writing here? Kind of. At least in terms of people who felt compelled
to complete the survey, I'm largely speaking to an audience of scientific
peers. BUT, I'm also reaching at least a few people who are not trained
scientists, and I suspect a lot more of the casual hits my blog receives each
day are not from dedicated scientists. Thanks again to everyone who
participated in the survey – many of you left awfully nice comments for me,
which really made me feel like this continues to be worth doing, so thanks for
the ego boost. It's been interesting to see who this blog is reaching and why
people are reading it! I don't know how long I'll keep this blog going, but I
don't have any intention of stopping soon and I'm glad I have an audience of
people who think it's worth following.

Sunday, December 6, 2015
Heroes in a half shell

In my continued quest to betray my dinosaurian research roots, I went to the American Museum of Natural History in New York to look at turtles! And what turtles they were - this is the skull of Ninjemys (the ninja turtle!), a giant meiolaniid turtle from Australia. Meiolaniids are the best turtles you've never heard of and it's a crying shame that they don't feature more prominently in prehistoric popular media.

Meiolaniids are basically ankylosaur-convergent turtles, all of which are very large for turtles and some, like Ninjemys, which would have been legitimately huge. They are only found in the southern hemisphere (and really mostly in Australia and nearby), and they all have elaborate cranial ornamentation. On the left in this picture is that same Ninjemys skull cast, and on the right we have Niolamia from Argentina - smaller, but arguably even weirder with those huge nuchal crests flaring off the back of the skull.

I'm interested in meiolaniids at the moment because they are one of the other groups of tetrapods that evolved a club-like tail weapon. Here's Meiolania platyceps, an island dwarf (!) meiolaniid, with some kind of boring mammal for scale.

And here's the business end of Meiolania. Stay tuned for more exciting discussion of animal business ends in the future!

Since we're looking at animal butts today, apparently, here are some Edmontosaurus rears...

...and a Triceratops derriere, for good measure.

Elsewhere in the fossil halls, I enjoyed meeting the disembodied floating head of this indricothere, which makes for a nice counterpoint to the fleshed-out model currently on exhibit at the North Carolina Museum of Natural Science's feature exhibit Extreme Mammals.

And here's a lovely trio of cingulates! The big fellow is the derived glyptodont Panochthus, the medium-sized fellow is the early glyptodont Propalaeohoplophorus, and off towards the back the little guy is the extant six-banded armadillo, Euphractus sexcinctus. The tail club on the Panochthus is mounted really weirdly, with it sticking straight out from the back of the shell rather than having any of the mobile rings from the base of the tail present, so I'm not sure what's quite up with that.

I post woefully few fish fossils on this blog, so here's a pretty great semionotid!

Science and anatomy and zoology are all around us if you keep your eyes open! I loved this chameleon logo on the side of a restaurant we came across one evening.
Subscribe to:
Posts (Atom)